



Nonvascular Plants

Although embryophytes are all decended from a shared land plant ancestor with a multicellular diploid sprophyte stage, being nonvascular is an ancestral trait inherited from green algal-like ancestors rather than a synapomorphy that can be used to show they are monophyletic. Presence/absence data of mitochondrial introns suggests they are not monophyletic, with Liverworts as sister to all other embryophytes (Qiu et al. 1998). A number of molecular phylogenies have corroborated this, showing a grade with Hornworts as sister to vascular plants (e.g. Chang and Graham 2011). These relationships are shown in the tree below.

More recently, however, a study using large amounts of nuclear transcriptome data (Wickett et al. 2014) suggests Bryophytes may actually be monophyletic as illustrated below.
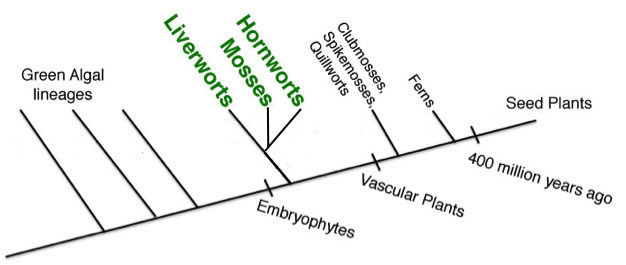
Some analyses in Wickett et al. 2014 put Hornworts by themselves as the first-branching lineage of Bryophytes. Given the conflicts in analyses, the phylogeny at the top of the page shows the relationships among bryophyte lineages and between bryophytes and vascular plants to be unresolved.
Nonvascular plants don't possess true vascular tissue (xylem and phloem), all have a long-lived gametophyte generation that produces gametes (sperm and egg), and all have a relatively short-lived diploid sporophyte generation that produces spores through meiosis, but these are all ancestral characteristics inherited from aquatic algal ancestors rather than shared, derived characters (synapomorphies) that would group them together as monophyletic.
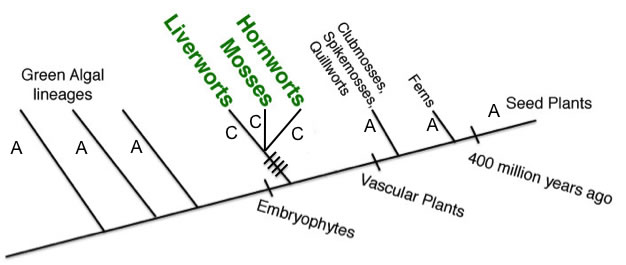
The following tree shows a DNA character that behaves in the same way as the morphological characters of no vasculature vs. vasculature or dominant haploid generation vs. dominant sporophyte generation. Having "A" at this nucleotide position is ancestral, a character found in algal outgroups as well as all nonvascular plants, so it does nothing to group the nonvascular plants together as monophyletic. The switch to a "C" at this nucleotide position can be inferred to have occurred on the branch leading to the common ancestor of all vascular plants in the same place that true xylem and phloem, as well as a dominant sporophyte generation, evolved as synapomorphies of the group.

By contrast, characters that support all nonvascular plants as a monophyletic group are shared across the bryophyte lineages but are not present in green algae or vascular plants. Characters of this type indicate changes that occurred along a branch leading to the common ancestor of nonvascular plants after it diverged from the vascular plant lineage but before divergence of Liverworts, Mosses, and Hornworts from each other. The tree below shows where synapomorphies of nonvascular plants occur which support monophyly of bryophytes.
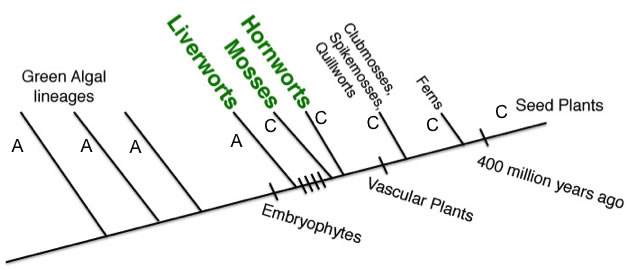
If Mosses and Hornworts are actually more closely related to vascular plants than they are to Liverworts, then we should see changes that link them to vascular plants at nucleotides where Liverworts retain the ancestral character state. These types of characters evolved on the branch leading to the common ancestor of these lineages after the branch leading to Liverworts had already diverged. The following tree shows a nucleotide character that supports Mosses and Hornworts being more closely related to vascular plants than they are to Liverworts (i.e. a synapomorphy of Mosses+Hornworts+Vascular Plants). In studies like the Qiu et al. 2008 and Chang & Graham 2011 papers linked above, there are more characters like this than those that support monophyletic nonvascular plants.
Marchantiophyta (Liverworts): 9,000 species
The sporophyte generation of most liverworts is very inconspicuous; although it is multicellular, like other embryophytes, it usually remains mostly hidden beneath the umbrella-like, egg-producing gametophores. Male gametophores may also be umbrella-shaped or may be patches of tissue on the surface of the thallus; they produce sperm on their upper surface so that raindrops landing on the gametophore can splash sperm away from the parent gametophyte. Gametophores are haploid tissue, just like the primary photosynthetic tissue of the liverwort, and produce sperm and egg cells with mitotic haploid nuclei identical to the cells in the plant body and gametophore. Thus, in liverworts, it is haploid tissue that forms the stalk of the structure that is responsible for lifting spores high enough to be dispersed away from the mother plant. In mosses, liverworts, and vascular plants, that job is performed by diploid sporophyte tissue. Liverworts are roughly divided into two groups: thalloid liverworts, which grow as thalloid mats of tissue undiferentiated into clear stem-like and leaf-like structures, and leafy liverworts, which are more common in the tropics and have small bits of leaf-like, photosythetic tissue attached to a more stem-like structure.
Reboulia hemisphaerica: gametophores and gametophytes, underside of gametophore showing sporophytes
Asterella tenella (gametophytes with gametophores)
Conocephalum conicum (air pores)
Marchantia polymorpha (gemmae and gemmae cups)
Thalloid Liverwort sp., Costa Rica: Gametophores and Gametophytes
Leafy Liverwort sp., Costa Rica: Gametophyte
Bryophyta (Mosses): 12,000 species
Mosses are the most species-rich lineage of nonvascular plants. They also have a disproportionate ecological impact, often colonizing new habitats before other embryophytes and forming a base organic substrate for other larger plants to colonize. Species of Sphagnum (peat moss) are particularly important in wet, low nutrient habitats. Many species of Sphagnum eventually form floating mats that can grow into islands or 'quaking bogs' which are colonized by grasses, sedges, carnivorous plants, and eventually shrubs and trees once the mats grow thick enough. Dried blocks of dead peat moss are used in specialized potting soils or as soil amendments and are also occasionally burned for fuel in subarctic regions where wood is scarce.
The photosynthetic 'cushion' portion of mosses is the gametophyte which makes egg-producing archegonia and sperm-producing antheridia. When the surface of the moss is coated with a film layer of water, the antheridia release swimming sperm which search out and fertilize the egg inside an archegonia of a different plant. The resulting zygote then grows into the sporophyte generation, which is often shaped like a golf club or stalked lantern. Both the stalk and the 'club' are formed from diploid tissue, and the diploid cells inside the sporangium at the top of stalk undergo meiosis to produce haploid spores. While the diploid generation in mosses is larger and longer-lived than in liverworts, it is still reliant upon the photosynthetic, haploid gametophyte out of which it grew for its nutritional needs.
Polytrichum commune Reindeer Moss, sporophytes and gametophytes
Moss sporophytes
Antheroceratophyta (Hornworts): 160 species
Hornworts are generally inconspicuous, with gametophytes resembling thalloid liverworts. The 'horn' is the diploid sporophyte generation, and splits violently in a corkscrew pattern when dry and mature to disperse the spores. The sporophyte generation is usually green, somewhat photosynthetic, and possesses stomata-like pores on its surface to aid in nutrient transport.Anthoceros sp. (from Costa Rica)




